端粒:细胞生命计时器与染色体保护机制
发布时间:2026-07-13
端粒可被理解为染色体末端的保护结构,类似于鞋带两端的塑料封头,其主要功能为保护遗传信息完整性并调控细胞分裂次数,因此亦被称为细胞的"生命计时器"。
在正常细胞分裂过程中,端粒随每次分裂而逐渐缩短。当端粒缩短至临界长度,染色体将失去有效保护,细胞随即进入分裂停滞状态,逐步走向衰老或死亡。这一过程既是个体衰老的重要驱动力,也是机体防止细胞异常增殖、抑制肿瘤发生的一道天然屏障。
然而,该屏障可被突破。当细胞获得维持端粒长度的能力时,即可实现无限分裂,转变为"永生性"细胞,这也是绝大多数恶性肿瘤得以持续生长的关键机制。
人类癌细胞主要通过两种方式实现端粒长度维持。约85%~90%的癌症通过激活端粒酶直接延长端粒;而其余10%~15%的端粒酶阴性肿瘤则依赖替代性机制维持端粒,即端粒延长替代机制(Alternative Lengthening of Telomeres, ALT)。ALT并非随机产生的异常现象,而是在端粒酶沉默且分裂压力显著增强的条件下,细胞启动DNA修复及同源重组过程,从而实现端粒的延长[1]。
以ALT机制维持永生性的肿瘤多源于间质组织,常见于肉瘤、神经胶质瘤及神经内分泌肿瘤等类型。此类肿瘤通常具有侵袭性强、易复发转移及临床预后较差的特点[2]。由于不依赖端粒酶活性,传统端粒酶靶向药物对ALT肿瘤基本无效,使其成为临床肿瘤治疗中极具挑战性的难题。
ALT与衰老的内在联系
近年研究发现,ALT与衰老并非彼此孤立的生物学过程,二者存在密切关联。端粒酶阴性的癌细胞与衰老细胞在多个表型上具有相似性,包括DNA复制过程持续受损、复制叉停滞频率升高;细胞内蛋白质修饰系统发生广泛改变,相关修饰介导的特殊结构易发生异常;细胞核内染色质的结构状态亦出现显著变化(图1)。上述特征提示,ALT阳性癌细胞与衰老细胞可能共享同一套调控逻辑。ALT甚至可被视为衰老细胞在规避死亡、转变为恶性细胞过程中的潜在路径之一。
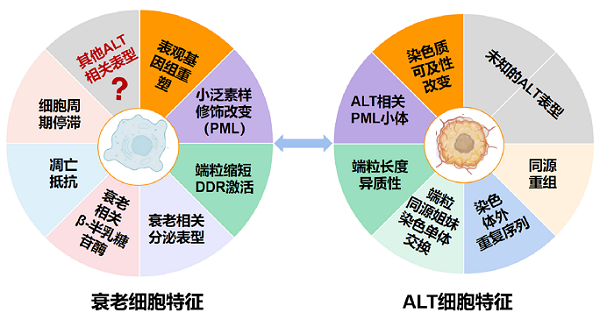
图1.衰老细胞和ALT细胞共性连接
激活ALT的癌细胞具有若干典型特征:端粒长度差异显著;形成特殊的APBs结构;端粒之间频繁发生片段交换;细胞内存在大量游离的端粒碎片。与之对应,正常衰老细胞同样表现为端粒酶失活、端粒持续缩短、DNA持续受损,部分衰老细胞亦可形成类似APBs的结构,并呈现染色质状态的显著改变[3]。由此可见,衰老与ALT肿瘤并非两条完全独立的生物学路径,而是在共同的"端粒危机"背景下,细胞选择的不同命运走向。
细胞命运的决定因素:p53基因的核心作用
决定细胞走向衰老还是癌变的关键调控因子为p53基因[4]。当p53功能正常时,端粒缩短至极限后,细胞将进入衰老状态,停止分裂,从而降低癌变风险。而当p53发生突变或功能丧失时,细胞失去增殖刹车机制,在持续压力下激活ALT,通过重组方式不断延长端粒,最终突破增殖限制,演变为恶性肿瘤[5]。此外,蛋白质修饰、染色质结构状态等细微差异亦可在"衰老"与"癌变"之间调控细胞命运,使两者的界限呈现模糊化趋势[6-7]。
衰老与肿瘤关系的重新审视
传统观点认为,细胞衰老是机体抵御肿瘤的重要保护机制。然而,最新研究提示,衰老细胞并非处于稳定终点状态。在端粒持续受损、基因突变等压力条件下,部分衰老细胞可能重新激活端粒维持机制,特别是ALT,从而突破衰老限制,转变为恶性肿瘤。这意味着,衰老并非不可逆的终点。在长期机体损耗及基因变化影响下,部分衰老细胞可突破原有束缚,演变为癌细胞。这一机制亦可解释为何年龄增长与癌症发病率升高之间存在密切关联。
基于端粒与ALT的协同干预策略
鉴于端粒及ALT在衰老与肿瘤发生发展中的核心作用,新的干预策略正从传统单一靶向模式转向协同调控方向。基于ALT将衰老与肿瘤相连接的理论基础,新型治疗思路包括:通过药物破坏ALT依赖的APBs结构,阻断端粒延长路径;利用ALT肿瘤对DNA修复机制的高度依赖性,应用特异性抑制剂精准杀伤该类癌细胞;同时清除衰老细胞与ALT肿瘤细胞,增强免疫监视功能[8-9]。上述策略不再将衰老与肿瘤割裂对待,而是以端粒稳态为核心,实现抗衰老与抗肿瘤的协同干预。
展 望
端粒通过ALT机制将衰老与肿瘤这两大生命科学核心问题紧密连接,不仅为揭示衰老本质提供了关键理论支撑,也为端粒酶阴性的难治性肿瘤开辟了全新的治疗方向。端粒的长度维持、结构稳定性及信号输出贯穿细胞正常生长、衰老与恶性转化的全过程,是调控生命进程的核心元件。深入解析端粒损伤、ALT激活与细胞命运转换之间的精密调控网络,有助于重新理解衰老与肿瘤的内在联系,推动开发兼顾抗衰老与抗肿瘤的新型干预手段,为延长健康寿命、攻克恶性肿瘤提供更为系统的理论基础与可行的技术路径。(详情请点击阅读原文)
参考文献
[1] Tsatsakis A, Oikonomopoulou T, Nikolouzakis T, et al. Role of telomere length in human carcinogenesis (Review). Int J Oncol, 2023, 63: 78
[2] MacKenzie D Jr, Watters A K, To J T, et al. ALT positivity in human cancers: prevalence and clinical insights. Cancers, 2021, 13(10): 2384
[3] O'Sullivan R J, Greenberg R A. Mechanisms of alternative lengthening of telomeres. Cold Spring Harb Perspect Biol, 2025, 17(1): a041690
[4] Kroemer G, Maier A B, Cuervo A M, et al. From geroscience to precision geromedicine: Understanding and managing aging. Cell, 2025, 188(8): 2043-2062
[5] Macha S J, Koneru B, Burrow T A, et al. Alternative lengthening of telomeres in cancer confers a vulnerability to reactivation of p53 function. Cancer Res, 2022, 82(18): 3345-3358
[6] Andreou A M, Tavernarakis N. Roles for SUMO modification during senescence. In Protein Metabolism and Homeostasis in Aging. Boston, MASpringer US2010: 160-171
[7] Bettin N, Vaurs M, Decottignies A. Epigenetics of human telomeres. Cold Spring Harb Perspect Biol, 2025, 17(11): a041706
[8] Gaela V M, Hsia H Y, Joseph N A, et al. Orphan nuclear receptors-induced ALT-associated PML bodies are targets for ALT inhibition. Nucleic Acids Res, 2024, 52(11): 6472-6489
[9] Ngoi N Y L, Silverman I M, Johnson A, et al. Exceptional response to the ATR inhibitor, camonsertib, in a patient with ALT+ metastatic melanoma. npj Precis Oncol, 2025, 9: 227
作者简介
刘 静:昆明理工大学基础医学院副教授;研究方向:衰老及衰老相关疾病的功能研究和活性小分子药物研发。
贾同欣:昆明理工大学基础医学院研究生;研究方向:衰老及抗肿瘤药物药理研究。
(作者:刘静、贾同欣)
(本文来源于公众号:生物化学与生物物理进展)
附件下载:
上一篇: