 最新报道
最新报道溶酶体是细胞内的物质降解、循环和信号中心,对细胞稳态调控、发育和衰老至关重要。溶酶体功能紊乱与多种疾病的发生发展密切相关。为了满足不同的生理需求,溶酶体通过不断的融合和分裂重塑其形态与功能。相比于融合过程,目前对于溶酶体分裂过程的了解非常有限,相关调控因子及作用机制仍不清楚,执行溶酶体膜分裂的分子尚未被揭示。
2024年3月27日,中国科学院生物物理研究所王晓晨研究组与冯巍研究组合作在《Nature》杂志在线发表了题为"The HEAT repeat protein HPO-27 is a lysosome fission factor"的文章。该研究发现了溶酶体膜分裂因子并揭示了其作用机制。
研究人员建立了以线虫为模式的多细胞生物溶酶体研究体系,通过非偏好性遗传筛选鉴定得到HEAT 重复蛋白HPO-27,与人类MROH1同源。HPO-27及MROH1含有37个HEAT重复序列,它们的功能尚未被解析。
结合活体成像追踪及冷冻电镜等实验手段,研究人员发现HPO-27在线虫多个发育阶段及多个组织中广泛表达。hpo-27功能缺失造成线虫表皮细胞中累积大量管状溶酶体,形成管状网络,并最终导致溶酶体结构损伤。溶酶体功能分析显示,HPO-27及MROH1功能缺陷导致溶酶体酸性异常、水解酶活性降低,降解能力减弱。hpo-27突变体线虫表现胚胎死亡、幼虫发育停滞和寿命缩短等严重缺陷。上述结果表明,HPO-27在溶酶体形态、结构及功能完整性,以及线虫正常生命活动中发挥重要作用。
研究人员在线虫及哺乳动物培养细胞中同步开展系列实验,探究了HPO-27和MROH1调控溶酶体形态及功能的作用机制。1)共定位分析显示,HPO-27及MROH1均与溶酶体存在共定位并富集在溶酶体膜的特定区域。溶酶体动态变化分析表明,HPO-27缺失导致溶酶体分裂事件减少,融合事件增加;过表达HPO-27则显著降低管状溶酶体的数量。与此一致,MROH1 KO细胞中,管状再生溶酶体数量增加,分裂减少。2)HPO-27和MROH1与小G蛋白RAB7在溶酶体上共定位,且RAB7活性丧失完全抑制HPO-27/MROH1在溶酶体的富集。体内与体外结合实验表明,HPO-27/MROH1与活性状态的、而非失活状态的RAB7存在直接相互作用,表明HPO-27/MROH1作为RAB7的效应子被招募到溶酶体。3)超分辨显微镜连续成像结果显示,HPO-27和MROH1被招募至溶酶体,富集在膜分裂位点,介导溶酶体管状膜的缢缩和裂解。上述结果表明,HPO-27和MROH1被RAB7募集至溶酶体,调控溶酶体膜分裂。
接下来,研究人员进一步探究了HPO-27和MROH1介导溶酶体膜分裂的具体机制。他们在昆虫细胞表达纯化HPO-27和MROH1蛋白,利用体外重构的膜分裂实验(SMrT assay)检测其是否具有膜分裂活性。研究人员发现,HPO-27和MROH1蛋白可结合磷脂酸(phosphatic acid,PA),在体外构建的含有PA及荧光染料的稳定支撑膜管(SMrTs)上发生富集,介导膜管的缢缩和断裂。结合结构预测、截短体构建、超分辨显微成像、负染电子显微镜机体外膜管分裂活性检测等方法,研究者发现,单体HPO-27或MROH1可通过首尾相接的形式发生寡聚化,在线虫、哺乳动物细胞以及体外重构系统中介导溶酶体及体外重构膜管的缢缩与分裂,而HPO-27的末端HEAT 重复结构域为其自组装、溶酶体富集、膜分裂活性所必需。
综上,该研究发现HPO-27及其同源蛋白MROH1是溶酶体膜分裂因子,其通过自组装介导溶酶体管状膜的缢缩和分裂。值得关注的是,与经典的膜分裂因子Dynamin及其超家族成员不同,HPO-27和MROH1不具有ATPase或GTPase活性。在体外膜分裂实验中,它们以不直接消耗能量分子(ATP/GTP)的方式介导膜管分裂,表明HPO-27和MROH1是一类新的膜分裂因子。HPO-27和MROH1的发现为溶酶体膜及其他膜分裂的研究提供了新途径,为进一步揭示溶酶体稳态调控及生理功能奠定了重要基础。
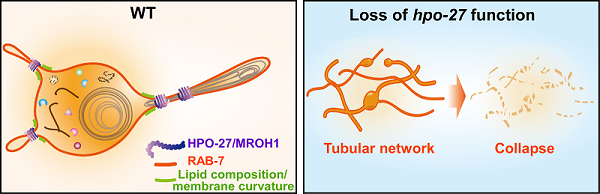
图:HPO-27介导管状溶酶体分裂的工作模型
中国科学院生物物理研究所王晓晨研究员和冯巍研究员为论文的共同通讯作者,云南大学和生物物理研究所联合培养博士生李乐涛,生物物理研究所刘希璐博士、博士生杨珊珊为共同第一作者。此外,生物物理研究所李栋研究员、胡俊杰研究员、武彦威博士、胡斯琪博士、张倩倩博士、张俊兵博士、胡俊岩博士、刘禹兵博士、王文娟博士、马晓丽博士、蒋阿敏博士,云南大学生命科学学院杨崇林教授也对该研究做出了重要贡献。该研究得到国家自然科学基金、国家重点研发计划、中国科学院战略性先导科技专项等项目的资助。
文章链接:https://www.nature.com/articles/s41586-024-07249-8
(供稿:王晓晨研究组)
 附件下载:
附件下载: