 最新图片新闻
最新图片新闻细胞焦亡作为机体重要的天然免疫反应,在拮抗和清除病原菌感染中发挥关键作用。当革兰氏阴性菌侵入宿主细胞后,其外膜的重要病原分子模式LPS(脂多糖,也称内毒素)会被宿主细胞内的天然免疫受体caspase-4/5/11识别,LPS激活的caspase-4/5/11会进一步切割活化焦亡蛋白GSDMD释放其膜打孔活性,导致细胞焦亡,激发宿主的抗菌炎症反应。同时,细菌也采用了多种策略来逃避宿主的免疫防御,例如通过独特的III型分泌系统向宿主细胞"注入"专门的效应蛋白,干扰宿主的免疫防御通路。2021年,北京生命科学研究所邵峰团队发现痢疾杆菌(Shigella)分泌的效应蛋白OspC3可以特异识别宿主细胞内的天然免疫受体caspase-4/11,通过催化caspase酶活中心的关键精氨酸发生一种全新的ADP-riboxanation翻译后修饰使caspase-4/11失活,阻断其活化下游GSDMD介导的细胞焦亡免疫防御。然而效应蛋白OspC3是如何特异地识别宿主靶标caspase-4/11,又是如何催化新颖的ADP-riboxanation修饰拮抗细胞焦亡的精确分子机理等关键科学问题有待进一步回答。
2023年1月9日,中科院生物物理研究所王大成/丁璟珒课题组和北生所邵峰团队合作,在《Nature Structural & Molecular Biology》发表题为"Structural mechanisms of calmodulin activation of Shigella effector OspC3 to ADP-riboxanate caspase-4/11 and block pyroptosis"的研究论文。该研究揭示了效应蛋白OspC3利用宿主细胞的钙调蛋白(calmodulin,CaM)作为辅助因子激活其酶学活性,特异地识别宿主靶标caspase-4/11并催化全新的精氨酸ADP-riboxanation修饰,阻断宿主细胞caspase-4/11-GSDMD焦亡通路的完整分子机理。
研究人员首先发现OspC3可以有效地对静息状态未发生自剪切的caspase-4/11和细菌LPS激活后发生了自剪切的caspase-4/11两种形式进行修饰,但是激活形式的caspase-4/11活性中心如果被模拟底物切割位点四肽序列的共价抑制剂zVAD不可逆地占据,会极大地削弱OspC3对caspase-4/11的修饰,这表明底物非结合状态的caspase-4/11,不论激活与否都是OspC3的底物。研究人员随后发现,ADP-核糖基特异性结合蛋白Af1521可与修饰后的caspase-4/11产物形成稳定的1:1复合物,通过解析Af1521与caspase-4被修饰后产物的复合物的高分率晶体结构,研究人员首次清晰地观察到ADP-riboxanation修饰的精确化学结构,caspase-4修饰位点R314的侧链胍基脱去一个末端Nω原子后与来自NAD+的ADP-核糖基核糖环上的C1原子和C2位羟基分别连接,形成了一个全新的五元恶唑烷环,该结果为精氨酸ADP-riboxanation这种全新的翻译后修饰提供了直接的结构证明。
OspC3及其所属的细菌效应蛋白家族具有典型的双结构域特征,其N端结构域和任何已知的蛋白质没有序列同源性,而C端包含一个保守的ankyrin-repeat结构域(ARD),这类结构域通常介导蛋白质相互作用,因此推测该结构域是OspC3的底物识别结构域。研究人员进一步通过解析OspC3 ARD结构域与caspase-4底物复合物的晶体结构,确定了OspC3 ARD结构域通过一系列氢键网络和疏水作用特异地招募宿主靶标caspase-4/11。
在体外重组OspC3修饰caspase-4/11的酶活实验中,研究人员发现OspC3需要几乎与底物蛋白caspase-4/11相当的量才能实现对底物的完全修饰,这有悖于酶催化底物反应高效性的经典认识。通过免疫共沉淀结合质谱的方法,研究人员鉴定出宿主的钙调蛋白CaM以Ca2+-free的形式与OspC3形成稳定的二元复合物,极大地提高了OspC3的催化效率。随后,研究人员成功地解析了OspC3与CaM二元复合物的高分辨率晶体结构,发现Ca2+未结合状态的CaM通过两个亚结构域分别以广泛的疏水作用牢牢抓住OspC3的N端结构域,而OspC3的N端结构域呈现出经典Rossmann折叠构象,与已知的ADP-核糖基转移酶结构域具有相似的结构特征和保守的NAD+结合基序。为了进一步阐明OspC3利用NAD+作为供体催化caspase-4/11的精氨酸发生ADP-riboxanation修饰的完整酶学机理,研究人员又成功解析了OspC3-CaM-caspase-4三元复合物及其与2'-F-NAD+(非水解型NAD+类似物)的四元复合物晶体结构,发现OspC3的N端的酶活中心通过酸性氨基酸D231固定caspase-4活性中心R314侧链胍基的末端Nω原子,使ADP-核糖基C1位靠近R314胍基的Nδ原子,从而利于NAD+烟酰胺基团的离去和在修饰位点R314 Nδ原子上发生第一步经典的ADP-核糖基修饰;而酶活中心的另一个酸性氨基酸D177则负责激活ADP-核糖基C2位的羟基亲核进攻R314侧链胍基C原子发生脱氨反应,使精氨酸侧链胍基和ADP-核糖基团形成一个恶唑烷环。随后,研究人员将结构研究的发现利用定点突变的方法在生化、细胞和Shigella感染小鼠三个层面进行了验证,完整地阐明了OspC3利用宿主辅因子CaM特异的对caspase-4/11进行精氨酸ADP-riboxanation修饰的分子机理。
这项研究工作通过一系列三维结构分析与功能实验验证,揭示了痢疾杆菌效应蛋白OspC3特异地识别宿主天然免疫受体caspase-4/11,并利用宿主钙调蛋白CaM作为辅因子催化全新的精氨酸ADP-riboxanation修饰,阻断宿主细胞caspase-4/11-GSDMD焦亡防御通路的完整分子机理,也为ADP-riboxanation这种全新的翻译后修饰的酶学反应机理提供了全面深入的理解,为进一步寻找和开发新型抗菌药物或细菌减毒疫苗提供了新策略。
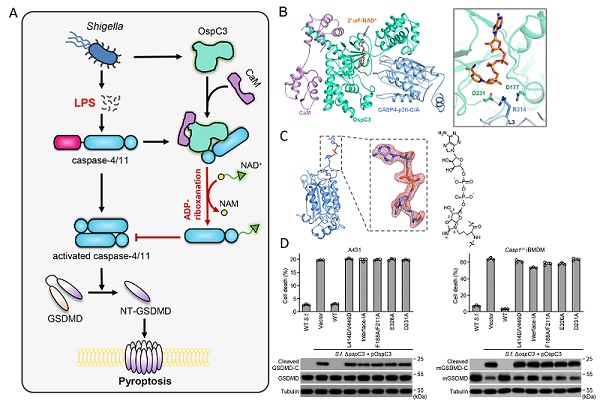
图 痢疾杆菌效应蛋白OspC3拮抗宿主细胞焦亡通路的分子机理
A. 痢疾杆菌效应蛋白OspC3阻断caspase-4/11焦亡通路拮抗宿主天然免疫的模式图;B. OspC3-CaM-caspase-4-2'-F-NAD+四元复合物结构及催化关键残基展示;C. 精氨酸ADP-riboxanation修饰后的caspase-4结构;D. OpsC3催化关键位点突变体在痢疾杆菌侵染细胞实验中进行验证。
中科院生物物理研究所丁璟珒研究员和北生所邵峰教授为本文的共同通讯作者,丁璟珒课题组侯彦婕博士和邵峰课题组生物物理所籍博士研究生曾欢为本论文的共同第一作者。该研究得到中科院战略先导科技专项、科技部重点研发计划、基金委优青项目及中科院青促会项目的支持。
文章链接:https://www.nature.com/articles/s41594-022-00888-3
(供稿:丁璟珒研究组)
 附件下载:
附件下载: