 2022年图片新闻
2022年图片新闻2022年4月7日,中国科学院生物物理研究所许瑞明研究团队在《Genes & Development》杂志在线发表了题为"Topography of histone H3-H4 interaction with the Hat1-Hat2 acetyltransferase complex"的文章,揭示了组蛋白乙酰转移酶Hat1-Hat2与完整组蛋白底物和组蛋白伴侣Asf1相互作用的结构基础,扩展了对组蛋白修饰酶与底物结合模式的认识,为理解组蛋白与多个组蛋白伴侣之间的动态结合机制提供了新的视角。
组蛋白翻译后修饰对于以染色质为基础的生命活动至关重要,涉及核小体组装、染色质复制、DNA损伤修复以及转录调控等过程。组蛋白乙酰化修饰是其中的一种重要类型,由组蛋白乙酰转移酶(HAT)负责催化完成。目前已经有十多种HATs被鉴定出来,单独的HAT或其与部分小肽底物的复合物结构也多有报道。但关于HAT-组蛋白相互作用的具体方式,尤其是涉及多个结构域或多蛋白形成的HAT复合物与完整组蛋白底物之间的催化和调节机制尚缺乏深入研究。
Hat1是最早被鉴定出来的组蛋白乙酰转移酶之一,可与组蛋白伴侣Hat2(RbAp46/48)一起组成Hat1-Hat2全酶复合体,催化新生组蛋白H4的第5位(H4K5)和第12位(H4K12)赖氨酸的乙酰化。本项工作中,研究组成功解析了酵母Hat1-Hat2与其完整组蛋白底物H3-H4、组蛋白伴侣Asf1以及辅因子CoA复合物的3.3 Å晶体结构,阐明了Hat1在Hat2辅助下催化组蛋白H4乙酰化的分子机制。结构显示,Hat1-Hat2结合完整H3-H4组蛋白底物的方式与结合组蛋白小肽的时候有很大差异,尤其是结构中组蛋白H4的α1螺旋回折与H3组成核心折叠区,修正了长期以来对H4与组蛋白伴侣Hat2/RbAP46/48结合方式的认知。该研究显示,组蛋白H3-H4核心折叠区结合在Hat1-Hat2形成的楔形区域,对于组蛋白异二聚体的稳定结合起到重要作用,有助于Hat1-Hat2高效乙酰化组蛋白H4。同时,结构中还观察到组蛋白H3的N端尾巴以一种舒展的构象与Hat2形成广泛的相互作用,这与研究组前期解析的另一个组蛋白乙酰转移酶Rtt109与底物H3-H4复合物结构(Cell, 2018)中观察到的H3状态相似,由此推测H3的这一区域可以采用不同的构象参与到多种生物学过程中。最后,结合研究团队近期对另一个组蛋白伴侣sNASP-组蛋白底物复合物的研究结果(Genes & Development, 2022),针对组蛋白H3尾巴在结合sNASP或Asf1时的不同构象,提出了细胞质中新生成的H3-H4经过一系列组蛋白伴侣的传递步骤,最终进入细胞核的动态模型。
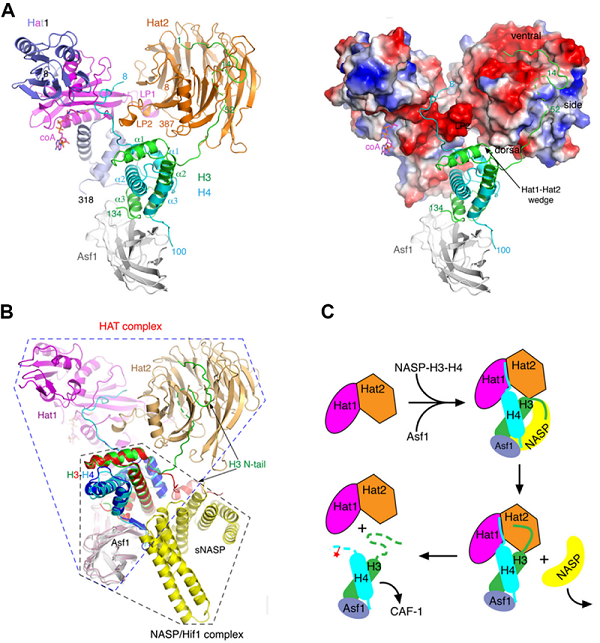
图1. Hat1-Hat2-Asf1-H3-H4-CoA复合物结构及其功能机理
A. Hat1-Hat2-Asf1-H3-H4-CoA复合物的晶体结构;B. 与Hif1/sNASP形成复合物的模型,该复合物由Asf1-H3-H4模块(蓝色虚线框内)与sNASP-H3-H4-Asf1结构(黑色虚线框内)叠加而成;C. H3-H4在组蛋白伴侣NASP/Hif1和Asf1之间传递的示意图。
中国科学院生物物理研究所许瑞明研究员为本文的通讯作者,博士研究生岳野和助理研究员杨文思为本文的共同第一作者,课题组博士生张林和项目研究员刘超培作出了重要贡献。该研究得到了国家自然科学基金、科技部重点研发计划、中国科学院战略性先导科技专项以及中国科学院青年创新促进会的经费支持。
文章链接:http://genesdev.cshlp.org/content/early/2022/04/05/gad.349099.121.short?rss=1
(供稿:许瑞明研究组)
 附件下载:
附件下载: