 2021年科研进展
2021年科研进展绿藻是水体和土壤中常见的光合生物,作为有机物的原初生产者在生态系统中发挥着重要的作用。它们具有和植物相似的两个光系统:光系统I(PSI)和光系统II(PSII),通过捕光复合物I和II(LHCI和LHCII)吸收光能并将能量传递给两个光系统,完成光驱动的电子传递和能量转换过程。在光照条件多变的自然环境中,光能在两个光系统之间的分配可能不均衡,这主要是由于PSI和PSII的捕光系统色素组成不同,导致它们对不同能量光子的吸收能力不同。
状态转换调控是光合生物适应光照条件变化、平衡激发能在两个光系统间分配的一种快速响应机制。当PSII被过度激发时,绿藻和植物PSII的捕光复合物LHCII被磷酸化,部分磷酸化LHCII三体与PSII解离并与PSI结合形成PSI-LHCI-LHCII超复合物,从而增加向PSI反应中心传递的能量,实现了激发能在PSII和PSI之间的平衡分配。
相比于植物,状态转换调控对于绿藻来说更为重要,绿藻中的PSI-LHCI-LHCII超复合物也比植物中相应的复合物更大更复杂,包含至少2个LHCII三体。在莱茵衣藻(一种单细胞模式绿藻)中,9个不同的LhcbM蛋白(LhcbM1-9,被分为4种类型)参与形成不同组成的LHCII三聚体。因而,莱茵衣藻PSI-LHCI-LHCII超复合物的结构和组成极为复杂,具体是由哪种LhcbM蛋白参与介导LHCII与PSI-LHCI复合物装配的问题一直以来都不清楚,其能量传递途径也有待通过高分辨率的结构研究来揭示。
生物物理所李梅课题组、柳振峰课题组联合日本国立基础生物学研究所Minagawa教授课题组,合作攻关并解析了莱茵衣藻PSI-LHCI-LHCII超复合物的高分辨率冷冻电镜结构。研究结果表明,该超分子复合体是由PSI核心在一侧结合8个LHCI蛋白,在另一侧结合2个LHCI单体和2个磷酸化LHCII三聚体,构成一个总分子量达~1100 kDa的膜蛋白-色素复合体(图a)。
基于高质量电镜密度图,研究人员准确指认了参与构成该超复合物的磷酸化LHCII中全部LhcbM蛋白,发现LhcbM1和LhcbM5通过它们的磷酸化氨基末端区域直接与PSI核心相互作用(图b-c),且LhcbM5还介导并稳定了PSI核心与LhcbM1之间的装配。此外,对分别缺失LhcbM1(ΔLhcbM1)和LhcbM5(ΔLhcbM5)的衣藻突变株的生化、功能和结构研究结果表明,只有LhcbM5对于超复合体的形成是必不可少的,ΔLhcbM5突变株的状态转换受到严重影响(图d-e)。在ΔLhcbM1突变体中,LhcbM3蛋白取代了LhcbM1与PSI结合,因而ΔLhcbM1突变体中仍然能够形成PSI-LHCI-LHCII超复合物。研究结果揭示了绿藻PSI和两种磷酸化LHCII之间的特异相互作用和潜在的激发能转移途径,为理解光合作用状态转换的分子机理和进化演变过程提供了新的见解。
上述研究成果于2021年7月8日发表在《Nature Plants》期刊,题为“Structural basis of LhcbM5-mediated state transitions in green algae”。中国科学院生物物理研究所李梅研究员、柳振峰研究员和日本国立基础生物学研究所Minagawa教授为论文的共同通讯作者,潘晓伟研究员(现工作单位首都师范大学)、Ryutaro Tokutsu博士和博士研究生李安节为该项工作的共同第一作者。该研究工作得到了科技部重点研发计划、国家自然科学基金、中国科学院B类先导专项和中国科学院前沿科学重点研究项目的共同资助。数据收集和样品分析等工作得到了生物物理所“生物成像中心”、生物物理所蛋白质科学研究平台等有关工作人员的大力支持和帮助。
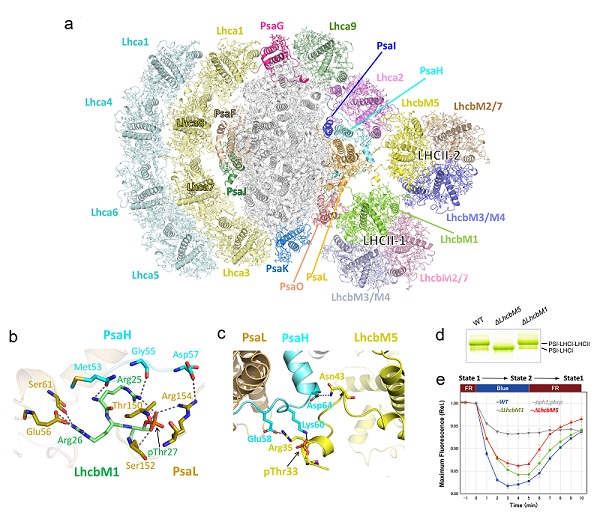
图:莱茵衣藻PSI-LHCI-LHCII复合物
文章链接:https://www.nature.com/articles/s41477-021-00960-8
(供稿:李梅研究组)
 附件下载:
附件下载: