高璞课题组和纽约大学合作
发布时间:2018-05-07
2018年5月3日,Cell杂志在线发表了高璞课题组与纽约大学合作的研究论文“Structural Insights into Non-canonical Ubiquitination Catalyzed by SidE”。该工作解析了来源于高致病性嗜肺军团菌(Legionella pneumophila)的新型泛素化酶SidE与ubiquitin和配体的多个复合物的高分辨率晶体结构,并结合大量的生化实验和突变体分析,揭示了SidE同ubiquitin及其配体的新颖互作方式和催化机制。
泛素化修饰调控着真核细胞几乎所有的生命活动。在与宿主的长期博弈过程中,病原菌进化出了多种方式来干预宿主的泛素修饰系统,从而达到增强侵染效率和逃避免疫监视的目的。以往报道的泛素化修饰(不论是宿主自身的,或是病原参与的),均是通过经典的E1-E2-E3三酶级联反应进行催化。而近期鉴定的来源于嗜肺军团菌的SidE家族却可以对多种宿主蛋白进行非典型的泛素化修饰。该家族有四个成员:SidE、SdeA、SdeB及SdeC,其结构域从N端到C端分别为DUB结构域、PDE结构域、mART结构域及coil-coil结构域。在经典的泛素修饰系统中,通过E1-E2-E3的级联反应,在ATP及Mg离子存在下,将ubiquitin的C末端共价连接到底物蛋白的赖氨酸上。而SidE家族蛋白参与的催化过程则完全不同:首先, mART结构域将NAD分子的ADP-ribose基团连接到ubiquitin第42位的精氨酸上,生成ADP-ribosylated ubiquitin(ADPr-Ub);然后,PDE结构域进一步切割ADPr-Ub中的磷酸二酯键,生成phospho-ribosylated ubiquitin(Pr-Ub),并同时将Pr-Ub共价连接到底物的丝氨酸上。然而,mART结构域结合及修饰ubiquitin,以及PDE结构域结合及水解ADPr-Ub的具体分子机制仍然不清楚。
在这项工作中,研究人员通过大量的尝试,成功获得了SidE在其两步催化反应中,与ubiquitin及其配体的高分辨率晶体结构。SidE的apo结构表明,mART结构域和PDE结构域的催化口袋互相远离并朝向不同的方向,提示这两个结构域的催化过程是相互独立的;同时这两个结构域又通过一段保守的基序紧密结合,当丧失这种相互作用时,两个结构域的活性都受到了影响。mART结构域与ubiquitin及辅因子NAD的复合物结构显示,mART结构域利用其表面的一个高度保守区域参与结合ubiquitin,而这个互作界面非常靠近mART的催化中心; ubiquitin的C末端tail则贡献了其主要的结合位点,尤其是第72位及74位的精氨酸紧密结合到mART结构域的酸性氨基酸形成的口袋里;NAD则结合于mART结构域经典的R-S-E motif上,其烟酰胺基团则朝向ubiquitin的42位精氨酸方向,等待着催化形成ADPr-Ub。PDE结构域与ubiquitin及ADP-ribose的复合物结构表明,ubiquitin结合于PDE结构域催化口袋的一侧,其Lys6-Thr9这段区域及His68贡献了主要的结合位点,Arg42朝向PDE结构域的活性中心;PDE结构域在结合了ubiquitin后会产生局部的构象变化,以利于催化的进行。以上所有参与结合及催化的关键氨基酸,研究人员均利用突变体实验进行了酶活验证。此外,研究人员还发现SidE家族蛋白对底物的识别不依赖于底物蛋白的特定三维结构,只要底物蛋白上含有能够进入SidE催化口袋的丝氨酸,那么均可被泛素化修饰。这些结果不仅较为完整的阐明了SidE家族蛋白新颖的泛素修饰机制,还为开发基于此新型泛素修饰系统的生物学工具提供了基础。
该工作是由中国科学院生物物理研究所高璞课题组与纽约大学合作完成的。高璞研究员为本文的主要通讯作者,高璞课题组助理研究员王勇博士为本文的第一作者。上海光源在晶体衍射数据收集过程中给予了帮助;生物物理所平台的杨福全研究员和谢振声老师在质谱分析方面给与了大力帮助;生物物理所俞洋研究员在细胞实验方面给予了指导。该工作得到了国家自然科学基金、中科院先导专项计划、生物物理所启动经费的资助。
文章链接:https://doi.org/10.1016/j.cell.2018.04.023
文章评论:高璞组揭示新型泛素化修饰的作用机制--胡荣贵、王丰点评
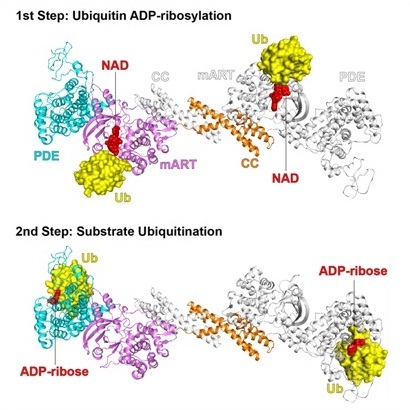
图一:SidE的两步反应中,mART结构域及PDE结构域结合ubiquitin的三维结构
(供稿:高璞课题组)
附件下载:
下一篇: