倾听百灵鸟的歌唱
发布时间:2020-04-07
语言在人类文明的发展中扮演着举足轻重的角色,人类语言在自然交际系统中似乎是独一无二的,甚至我们的近亲长臂猿或者黑猩猩等非人类灵长类动物也不具备语言学习的能力[1]。然而,却惊奇的发现鸟类表现出类似于人类语言学习的行为特征。目前已知人类语言的基础是发声学习。发声学习是一种通过模仿而获得发声的能力。你可能会问,很多动物也能发声,如鸡鸭狗,为什么鸟类的发声就与众不同呢?那是因为鸡鸭狗这些动物的发声是其本能获得的,它一般是由遗传决定种族特有的声音,比较简单。当然鸟类也有其特有辨识度的声音,我们把这种声音称为鸣叫,而我们要讨论的发声学习是通过感知记忆到内化后的发声,通俗的说就是我们在早期通过聆听和模仿成人(成年鸣禽)的声音,通过听觉反馈不断纠正自己发声的过程,我们把这种声音称为鸣唱。
鸣禽学习发声非常像人类婴儿获取口语。两者学习发声的过程在行为、神经和遗传上表现出许多惊人的相似之处[2]。
和人类一样,鸣禽同样拥有对发声学习极为敏感的关键期[3]。在发育的敏感期,人类婴儿和鸣禽模仿发声依赖于听觉引导,从"听"到音符传入大脑留下痕迹,才能学习记忆和声音输出[4]。假如在敏感期听觉功能损伤,与人一样,将引起严重的发声学习障碍。而在成年后听觉损伤,则会导致已经形成的稳定语言结构发生很大的退化,这些结果均表明听觉反馈对发声学习的过程及声音结构的稳定是至关重要的[5]。
鸣禽的鸣唱主要依赖于控制鸣禽发声学习的鸣唱控制系统。该系统由控制鸣曲可塑性的前端脑通路(anterior forebrain pathway)和保持鸣曲稳定性的发声运动通路(vocal motor pathway)相互调节控制发声学习[6-7]。令人惊奇的是,这些与发声学习相关的脑区与人类语言学习相关的脑区在解剖位置上达到高度的吻合。(图1)[8]。更令人兴奋的是,负责组织和产生语句的布洛卡区(Broca's area)在绝大多数人大脑中位于左半脑,而在鸟类中也发现了鸣唱控制的左侧半球优势现象[1],这在人类以外的其他动物中尚未发现。
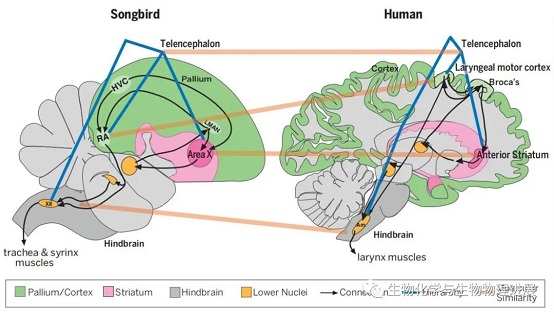
橙色区域为人类和鸟类拥有的功能相近的核团
另一个有趣的发现是鸟鸣作为鸟际间交流的语言和人类一样也具有地区性,俗称"方言"。主要表现为同种异歌现象。不同国度的同种鸟相会,会有明显的语言"隔阂",如百灵鸟的歌声,就有明显的地域之分[9]。鸣禽与人类这许多相似之处,使它们成为研究语音通信信号学习和处理学习、记忆与感知的重要模型。

不过,不是所有的鸟类都具备发声学习的能力,到目前为止,只有三个远亲鸟类(鹦鹉、蜂鸟和鸣禽)发现了这一种罕见的特征[10]。很多鸟类不仅可以学习本族的鸣曲,还能通过模仿其他鸟族的鸣声甚至人语,被我们广为熟知的要属鹦鹉了,这种能力称为效鸣。其同样具有发声学习的敏感期,在幼年时期模仿能力最强,曾有研究者给开始学唱的幼鸟如灰雀、乌鸫、欧椋鸟等,听管乐的乐曲声,结果它们就能发出管乐器的鸣声[9]。更有甚者,从幼年一直听其他鸣曲的鸟会放弃其种群的鸣曲,但当它听到种族的鸣声,能在很短时间内学会而改用本族的鸣曲。
而模仿力比较强的一些鸣禽,例如八哥、鹩哥等,能根据时间、地点、环境做出不同的反应。其中"鹦鹉学舌"是我们广为熟知的,当一只鹦鹉听到敲门声时,会突然大喊"请进来";当听到敲击木板声时,也会大叫"请进来"。你以为鹦鹉学会了人类的语言了吗?其实它们的这些反应都是无意识产生的,鹦鹉无意中把人们的敲门声和说"请进来"的两种声音联系起来,并把鸟脑受到的这种刺激信号贮存起来,形成了条件反射的缘故。但这种声音反射会令我们惊讶不已。

鸟类鸣唱的另一个奇特的现象是通常雄鸟善鸣而雌鸟很少或几乎不叫,且春季的雄鸟比秋季的更爱叫,这是因为雄鸟体内的雄激素水平高于雌鸟[11]、而春季又高于秋季的缘故。语言学家乔姆斯基认为:"只有假设人类先天便具备专门为学习语言而定制的神经组织,我们才有可能解释婴幼儿令人惊叹的语言学习能力。"通俗的讲就是结构决定功能,只有了解其神经生理和解剖结构,才能更透彻得了解人类语言的学习机制。《生物化学与生物物理进展》杂志(PIBB)2020年第一期发表的题为"雄激素调控鸣禽鸣唱核团对鸣唱行为影响"一文,根据作者描述雄激素可以通过调控细胞、电生理特性、突触、受体来影响鸣唱控制核团进而导致鸣唱行为改变。(详情请点击阅读原文)虽然鸟鸣声和人类言语的某些特征不是相似的,比如语言的意义能力、抽象能力和灵活的联想能力,但在感官体验如何内化并用于塑造声音的输出,以及如何在敏感期加强学习方面,有着惊人的相似之处。因此,可能涉及到类似的神经机制。这是我们不得不重视的,而且鸟鸣学习也被视为与人类语言学习最为接近的动物。
参考文献:
【1】Prather J F, Okanoya K, Bolhuis J J. Brains for birds and babies: Neural parallels between birdsong and speech acquisition. Neuroscience & Biobehavioral Reviews, 2017, 81(Pt B)
【2】Bolhuis J J, Okanoya K, Scharff C. Twitter evolution: converging mechanisms in birdsong and human speech. Nature Reviews Neuroscience, 2010, 11(11):747-759
【3】Aamodt C M, Farias-Virgens M. Birdsong as a window into language origins and evolutionary neuroscience.Biol. Sci,2020, 06:375(1789)
【4】Bolhuis J J, Moorman S. Birdsong memory and the brain: In search of the template. Neuroscience & Biobehavioral Reviews, 2015, 50(Sp. Iss. SI):41-55
【5】Zhou X, Fu X, Lin C, et al. Remodeling of dendritic spines in the avian vocal motor cortex following deafening depends on the basal ganglia circuit. Cerebral Cortex, 2017, 27(5): 2820-2830
【6】Goldberg J H, Fee M S. Vocal babbling in songbirds requires the basal ganglia-recipient motor thalamus but not the basal ganglia. Journal of Neurophysiology, 2011, 105(6): 2729-2739
【7】Tanaka M, Singh Alvarado J, Murugan M, et al. Focal expression of mutant huntingtin in the songbird basal ganglia disrupts cortico-basal ganglia networks and vocal sequences. Proc Natl Acad Sci USA, 2016, 113(12): E1720-E1727
【8】Pfenning A R, Hara E, Whitney O, et al. Convergent transcriptional specializations in the brains of humans and song-learning birds. Science, 2014, 346(6215):1256846
【9】李东风. http://blog.sciencenet.cn/blog-729911-1211993.html
【10】Jarvis E D. Learned birdsong and the neurobiology of human language. Ann N Y Acad Sci, 2004, 1016:749-777
【11】 Wang S, Liao C, Li F, et al. Castration modulates singing patterns and electrophysiological properties of RA projection neurons in adult male zebra finches. PeerJ, 2014, 2:e352
(作者:王青沁)
(本文来源于公众号:生物化学与生物物理进展)
附件下载:
上一篇:
下一篇: